第一部分RasGRP3负调TLRs触发的IL-6的产生及其分子机制研究
1引言
机体免疫系统能通过表达各类模式识别受体(pattern recognition receptors)来监视病原微生物的入侵并触发抗病原微生物的免疫应答Toll样受体(TLRs)是树突状细胞以及巨嗟细胞等天然免疫细胞上表达的一类至关重要的模式识别受体[2]。TLRs能有效识别来自病原微生物上的各类病原体相关分子并能触发MAPK(mitogen-activated protein kinases)通路和 NF-kB (nuclear factor kB)通路的活化’从而介导天然免疫细胞产生合适强度的抗感染免疫应答[3]。TLRs信号所触发的合适的免疫应答强度将有利于机体对入侵病原微生物的清除,同时也有利于机体维持正常的生理稳态;过强或过弱的免疫应答则会导致自身免疫性疾病的发生或导致病原微生物清除不完全而引发持续感染。目前对TLRs所介导的信号通路已经有了比较清楚的认识,即TLRs信号能通过MyD88依赖和非依赖两种途径进行传递。然而,目前对TLRs信号的调节,特别是对TLRs信号所触发的免疫应答强度的精细调节,仍不是非常清楚。ERK1/2被认为是MAPK中的重要成员之一。在TLRs配体作用下,ERK1/2会迅速发生活化[6]目前的研究表明ERK1/2活化后并不直接影响TLRs所介导的信号通路,而是影响了 TLRs所触发的免疫应答强度。
小GTP酵Ras被认为是一种分子开关,它能通过Ras/RafTMEK/ERK通路控制ERK1/2的活化[7-9]。在细胞中,Ras能以两种不同的形式存在,一种是与GDP相结合的非活性形式,另一种是与GTP相结合的活性形式[1°]。当细胞处于静息状态时,Ras主要以非活性形式存在;而当细胞受到刺激时,Ras则主要以活性形式存在,从而使其下游ERK1/2发生活化[11]。Rap也是小GTP酶家族中的一员,并携带有与Ras相同源的功能结构域。不过Rap对ERK1/2的活化作用目前还存在争议[m6],可能是因为不同的研究选择的细胞不同或使用的研究方法不一样,从而得出不一样的结论。在巨唾细胞中,特别是在TLRs所触发的信号中,Rap的功能还未见报道过。小GTP酶Ras或Rap的活性通常受到两类分子的影响:鸟核苷酸交换因子(GEF)和GTP酶活化蛋白(GAP)[i7,,GEF能促进GTP和GDP发生交换,导致Ras或Rap由非活性形式转变为活性形式;GAP能增强Ras或Rap本身的GTP酶活性,促进GTP水解为GDP,从而使Ras或Rap由活性形式转变为非活性形式。RasGRP家族成员是一类主要在淋巴细胞中高表达,并发挥重要功能的GfiF。RasGRPl能在T细胞中高表达,主要参与TCR信号的传递RasGRP2能在血小板中高表达,主要在凝血过程中发挥重要作用[21,22]; RasGRP3能在B细胞中高表达,主要参与BCR信号的传递RasGRP4能在肥大细胞中高表达,主要参与哮瑞等过敏反应[25,26]。
……….
2材料与方法
2.1实验材料
2.1.1实验动物
C57BL/6J(H-2Kb)小鼠,雄性,6-8周龄,购于上海必凯实验动物有限公司。RasGRPS转基因小鼠同野生塑C57BL/6J小鼠进行杂交,所得后代中杂合子和野生型小鼠用于相关实验分析。所有小鼠均于SPF级环境飼养,繁殖。
2.1.2细胞系人胚肾细胞系
HEK293和小鼠巨唾细胞系RAW264.7购自American TypeTissue Culture Collection,本实验室保存,传代。这两种细胞都用含10%胎牛血清的DMEM培养基培养。
2.1.3病毒
Herpes simplex virus I (HSV-1)由中国医学科学院医学生物学研究所李琦涵研究员惠赠。Vesicular stomatitis virus (VSV)由第二军医大学病原微生物教研室潘卫教授惠赠。Sendaivims(SeV)由本实验室护"增,保存。
2.1.4载体、菌株和分子克隆试剂
pcDNA3.1/His (-B)载体购自Invitrogen公司。DH5a感受态细胞购自北京全式金生物技术有限公司。细胞总RNA快速提取试别盒购自飞捷生物有限公司。反转录酶购自TOYOBO。高保真DNA聚合酵,限制性内切酶和DNA连接酶都来自TAKARA。琼脂糖DNA回收试劍盒,PCR产物纯化试剂盒以及普通质粒提取和纯化试刻盒购自上海生工生物工程有限公司。PCR引物由上海生工生物工程有限公司合成。琼脂糖购自华舜生物工程公司。
……….
2.1.5主要试剂和抗体
RPMI-1640、DMEM培养基和胎牛血清都购自PAA公司。LPS、G418、PD98059、Wortmannin、Go6983、弗氏完全佐刺和不完全佐剖、抗p-actin抗体和抗Flag标签表位抗体均购自Sigma-Aldrich公司。poly (I:C)购自Calbiochem,CpG ODN 购自 invivogen。抗 RasGRP、抗 ERKl/2、抗填酸化 ERKl/2 (202 位苏氨酸/204位路氣酸)、抗AKT、抗填酸化AKT(473位丝氣酸)、抗碟酸化AKT(308位苏氣酸)、抗JNK、抗碟酸化JNK1/2 (183位苏氧酸/185位赂氣酸)、抗P38、抗碟酸化P38 (180位苏氣酸/182位酪氧酸)、抗IKKa/p、抗碟酸化IKKa/p (176/180位丝氣酸)、抗IKBa和抗碟酸化IKBa (42位路氧酸)抗体购自Cell SignalingTechnology。抗碟酸化RasGRP (133位苏氨酸)购自Epitomics公司。质粒转染试刻JetPEI和siRNA转染试剂INTERFERin购自Polyplus公司。siRNA由上海吉玛制药技术有限公司设计并合成。Ras及Rapl活性分析试刻盒购自MilUpore公司。IL-6 ELISA试刻盒购自R&D公司。鸡11型胶原购自Chondrex公司。
2.1.6主要设备及软件普通
PCR仪和化学发光成像仪购自Gene公司,琼脂糖凝胶电泳仪购自天能公司,焚光定量PCR仪购自Roche公司,Western blot全套电泳转膜仪器和酶标仪均购自Bio-Rad公司,常温和低温高速离心机购自Eppendorf公司。引物使用Primer Premier 5.0软件进行设计。数据用GraphPad Prism 5统计软件进行分析。测序结果用Vector NTI Suite 7软件进行出对。
……….
3 试验结果...........49
3.1巨噬细胞能有效区分刺激物强度的细微差别......... 49
3.2作为第二信使的钙离子能反映刺激物强度的细微差别.........52
3.3 STIM1通过影响钙离子通道的开关来控制 .........58
3.4钙离子能协调小GTP酶RAS和RapI的活化强度......... 60
3.5钙离子能加速AKT去磷酸化并促进ERK活化.........62
3.6钙离子能影响感染局部的天然免疫应答......... 64
4讨论 .........66
5结论.........69
4讨论
TLRs信号通路的调控对于机体有效清除病原微生物以及维持正常的生理稳态非常重要;不当的TLRs信号强度容易导致不足或过强的免疫应答并容易引发病原微生物的持续感染或引发自身免疫性疾病的发生[46,47]。近年来,研究人员对TLRs的反馈调节巳经有了比较清楚的认识。然而,对TLRs的前馈调节却知之甚少。我们的研究发现作为第二信使的毎离子能在细胞内反映胞外TLRs刺激物强度的细微差别,并能向下游传递具有细微差别的信号,从而精细地调控巨瘦细胞做出与TLRs刺激物强度相匹配的应答。生理学上关于前馈调节的描述是:机体在发出一个动作电位指令的同时,会通过快捷方式发出另一条前馈调节指令,从而更准确地指导某个细微动作的完成。因此,从该意义上来讲,甸离子所介导的对TLRs所触发的免疫应答的调节,类似于一种对TLRs信号的前馈调节。在本课题研究的开始阶段,我们提出了一个假设,即TLRs在质膜上大致呈均句分布,不同强度的TLRs配体刺激时,能有效捕捉到配体的TLRs数量是不一样的,因此,TLRs能感知并能向胞内传递刺激物强度的差异信号,从而指导巨嗟细胞做出与激物强度相匹配的应答。为了证实这个假设,有一个关键问题需要解决。即剌激物强度的差异信息在如胞内如何得到反映并向下游传递。
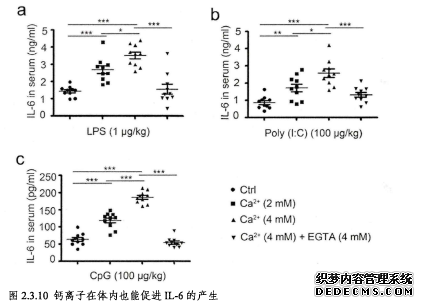
………..
结论
本部分的研究主要是在论文第一部分内容的提示作用下,继续探讨巨嗟細胞是如何感知刺激物强度的细微差别,并准确地做出与刺激物强度相四配的应答。
(1)巨嗟细胞能识别TLRs刺激物强度的微弱差别并能做出相应强度的应答;
(2)作为第二信使的每离子能反映TLRs刺激物强度的细微差别并介导巨唾细胞做出与刺激物强度相匹配的应答;
(3) STIM1通过影响甸离子通道的开关来控制成为第二信使的甸离子内流量,从而间接调节巨唾细胞的应答强度;
(4)锦离子能协调小GTP酶Ras和Rapl的活化强度;
(5)韩离子能加速AKT去碟酸化并促进ERK活化;
(6)甸离子能影响感染局部的天然免疫应答。
…………
参考文献(略)